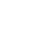清華新聞網(wǎng)9月9日電 新冠病毒自2019年12月以來蔓延至全球各地,特別是近期受到廣泛關(guān)注的德爾塔變種,平均能夠傳染8-9個(gè)人,傳播能力大幅度提高。新冠病毒的變種為什么能夠具有這么強(qiáng)的傳染能力呢?為了解釋這一原因,清華大學(xué)藥學(xué)院譚旭實(shí)驗(yàn)室和中國科學(xué)技術(shù)大學(xué)生命科學(xué)學(xué)院金騰川實(shí)驗(yàn)室從細(xì)胞免疫的角度出發(fā),通過高通量篩選的方法鑒定了新冠病毒特異性CD8+ T 細(xì)胞的抗原識(shí)別表位,并找到了新冠病毒變種逃逸細(xì)胞免疫的證據(jù),為后續(xù)疫苗的設(shè)計(jì)和優(yōu)化提供了新的思路。
近日,研究團(tuán)隊(duì)在國際一流期刊《細(xì)胞報(bào)道》(Cell Reports)上發(fā)表了題為“鑒定新冠康復(fù)者的細(xì)胞免疫表位發(fā)現(xiàn)新冠病毒變種的細(xì)胞免疫減弱的現(xiàn)象”(Profiling CD8+ T Cell Epitopes of COVID-19 Convalescents Reveals Reduced Cellular Immune Responses to SARS-CoV-2 Variants)的研究論文。

圖1:新冠病毒特異性CD8+ T細(xì)胞抗原表位的鑒定
新冠病毒德爾塔變種的命名來自于美國疾病控制中心和世界衛(wèi)生組織的分類系統(tǒng),由于新冠病毒在傳播過程中在不斷地變異,為了更好地定義新冠病毒的變種,美國疾病控制中心和世界衛(wèi)生組織獨(dú)立建立了一個(gè)分類系統(tǒng),將新冠病毒的變異體分為關(guān)注變種(Variants of concern)和待關(guān)注變種(Variants of interest)。關(guān)注變種的特點(diǎn)是:具有更高的傳染性,即可以感染更多的人群;誘發(fā)更嚴(yán)重的疾病 (即住院或死亡人數(shù)增加);降低治療和疫苗的有效性;可能逃避診斷檢測。
關(guān)注變種一共包括四種變異體:阿爾法變種 (B.1.1.7),貝塔變種 (B.1.351),伽馬變種 (P.1),德爾塔變種 (B.1.617.2) ;待關(guān)注變種被認(rèn)為是危險(xiǎn)性較低的關(guān)注變種,世衛(wèi)組織于2021年6月22日更新了待關(guān)注變種的名單,包括7個(gè)變異體:Epsilon (B.1.427和B.1.429),Zeta (P.2), Eta(B.1.525),Xita (P.3),Iota (B.1.526),Kappa (B.1.617.1)和最近廣受關(guān)注的拉姆達(dá)(C.37)。
全球流感共享數(shù)據(jù)庫(GISAID)總結(jié)了2021年所有在官網(wǎng)提交的新冠病毒序列的比例匯總(圖2),可以看到新冠病毒變種在全球感染范圍的動(dòng)態(tài)變化。

圖2:全球流感共享數(shù)據(jù)庫(GISAID)官網(wǎng)收錄的新冠病毒序列比例統(tǒng)計(jì)
從圖2可以看到,2021年1月起,阿爾法變種首先占據(jù)傳播優(yōu)勢。然而從5月開始,德爾塔變種逐漸代替阿爾法變種,成為在全球傳播的絕對(duì)優(yōu)勢變種。德爾塔變種是在2020年10月首次在印度馬哈拉施特拉邦被發(fā)現(xiàn)的。隨后在2020年末僅用2個(gè)月的時(shí)間傳播到整個(gè)印度,并超越了在印度傳播的原有的73個(gè)譜系,可見傳播能力之強(qiáng)。至今德爾塔變種已經(jīng)傳播到了200多個(gè)國家,是目前最值得關(guān)注的變種。
新冠病毒變種的優(yōu)勢建立和病毒的感染能力、免疫逃逸能力、傳播能力等方面都有關(guān)系。以德爾塔變種為例,已有研究報(bào)道,與阿爾法變種相比,德爾塔變種的S蛋白能夠更加高效地被宿主細(xì)胞蛋白酶切割形成具有功能的蛋白,在類器官水平上極大提高了德爾塔變種入侵細(xì)胞的效率,也加快了德爾塔變種在感染初期的復(fù)制速度,為德爾塔變種優(yōu)勢建立提供了思路。除了感染能力的提高,關(guān)注變種也增強(qiáng)了免疫逃逸能力,以德爾塔變種為例,和野生型新冠病毒相比,新冠病毒康復(fù)者血清中和抗體對(duì)德爾塔變種的中和能力平均下降6倍;疫苗(ChAdOx-1和BNT162b2)對(duì)德爾塔變種的防護(hù)效果平均下降8倍;臨床批準(zhǔn)的用于治療新冠病毒相關(guān)疾病的單克隆抗體bamlavinimab和imdevimab對(duì)德爾塔變種的中和能力分別下降1000倍和50倍。
雖然關(guān)注變種對(duì)抗體介導(dǎo)的體液免疫敏感度下降,但由于關(guān)注變種在傳播過程中絕大部分人都是初次感染,體內(nèi)并沒有中和抗體,因此對(duì)已知抗體的反應(yīng)較弱能夠提示我們現(xiàn)有疫苗對(duì)于這些變種的保護(hù)性會(huì)有所下降,但不能完全解釋關(guān)注變種的大規(guī)模傳播。此外根據(jù)實(shí)驗(yàn)數(shù)據(jù),德爾塔變種對(duì)康復(fù)血清或疫苗后血清的中和抗體敏感性下降程度和貝塔變種(B.1.315)接近,但傳播程度卻遠(yuǎn)超貝塔變種,這提示除了體液免疫之外,CD8+ T細(xì)胞介導(dǎo)的細(xì)胞免疫應(yīng)答的逃逸也可能促進(jìn)了新冠病毒優(yōu)勢變種的不斷傳播。先前的研究已經(jīng)表明,細(xì)胞免疫應(yīng)答不僅與新冠病情的輕重發(fā)展具有相關(guān)性,特別地,新冠病毒特異性CD8+ T細(xì)胞早于中和抗體被檢測到,說明細(xì)胞免疫應(yīng)答在病毒感染早期就開始發(fā)揮作用,而這種早期的免疫抑制對(duì)于機(jī)體防御新冠病毒、抑制新冠病毒的傳播可能是非常重要的。此外,對(duì)2003年SARS康復(fù)患者的長期研究表明,細(xì)胞免疫保持的時(shí)間長度很長,有的甚至能長達(dá)17年,相對(duì)而言,康復(fù)患者的抗體免疫記憶則要短很多。系統(tǒng)地了解新冠病毒感染者特別是康復(fù)患者的細(xì)胞免疫應(yīng)答,對(duì)于我們了解病人的病情發(fā)展的免疫決定因素,幫助新型疫苗設(shè)計(jì)甚至預(yù)測病人對(duì)疫苗的反應(yīng),都有重要意義。
探究細(xì)胞免疫應(yīng)答的重要步驟就是鑒定新冠病毒特異性CD8+ T細(xì)胞的抗原識(shí)別表位。清華大學(xué)譚旭課題組運(yùn)用T-Scan技術(shù),研究了能夠引起人體CD8 T細(xì)胞響應(yīng)的新冠病毒抗原表位的分布,并發(fā)現(xiàn)了新冠基因組上的4個(gè)氨基酸位點(diǎn)的突變可能賦予關(guān)注變種逃逸細(xì)胞免疫的能力從而促進(jìn)病毒的傳播,為關(guān)注變種的防范提供了細(xì)胞免疫方向的基礎(chǔ) (圖1)。
T-Scan技術(shù)是2019年由美國哈佛大學(xué)醫(yī)學(xué)院斯蒂芬·埃利奇(Stephen Elledge)教授發(fā)明的用于識(shí)別T細(xì)胞抗原的高通量篩選平臺(tái)。運(yùn)用T-Scan技術(shù)構(gòu)建新冠病毒抗原呈遞的靶細(xì)胞文庫的原理(如圖1所示)。首先運(yùn)用DNA芯片大規(guī)模合成技術(shù),合成覆蓋新冠病毒編碼的所有蛋白的多肽基因序列,每個(gè)多肽56氨基酸,一共有9179條序列。之后使用慢病毒載體將新冠病毒多肽抗原的基因文庫轉(zhuǎn)達(dá)到靶細(xì)胞中,在靶細(xì)胞內(nèi)進(jìn)行多肽抗原基因的表達(dá)和多肽抗原的內(nèi)源性加工最終由靶細(xì)胞的主要組織相容性復(fù)合體(MHC)分子將新冠病毒抗原呈遞到細(xì)胞表面,較好地模擬了細(xì)胞被病毒感染后將病毒抗原呈遞到細(xì)胞表面的過程。此外,靶細(xì)胞還攜帶響應(yīng)顆粒酶B(Granzyme B, GzB)活性的IFP熒光報(bào)告基因(圖3),用于篩選被CD8+ T細(xì)胞識(shí)別的靶細(xì)胞。GzB是一種絲氨酸蛋白酶,平時(shí)儲(chǔ)存在CD8+ T細(xì)胞內(nèi)的細(xì)胞毒性顆粒中,當(dāng)CD8+ T細(xì)胞識(shí)別靶細(xì)胞呈遞的病毒抗原后被活化,CD8+ T開始執(zhí)行殺傷功能,通過穿孔素在細(xì)胞膜上形成孔洞將裝載GzB的細(xì)胞毒性顆粒特異性分泌到靶細(xì)胞中。GzB進(jìn)入靶細(xì)胞后激活I(lǐng)FP熒光報(bào)告基因。接下來可以通過流式篩選富集被識(shí)別的靶細(xì)胞,并通過下一代測序鑒定靶細(xì)胞呈遞的抗原序列。

圖3: 響應(yīng)Granzyme B (GzB) 的熒光報(bào)告系統(tǒng)
CD8+ T細(xì)胞的識(shí)別具有MHC-I類分子限制性,只能識(shí)別和自身MHC基因型匹配的MHC分子所呈遞的抗原。MHC-I類分子包括A,B,C三種基因型,稱為HLA-A、HLA-B和HLA-C,每一種基因型又包括了多種等位基因,因此MHC分子的多樣性非常豐富。作者選取了中國人群中頻率最高的四種HLA-A基因型進(jìn)行研究,包括HLA-A*02:01、HLA-A*02:07、HLA-A*11:01和HLA-A*24:02(分別以占中國人口的10.9%、9.2% 、21.4%和14.4%)。
作者構(gòu)建了過表達(dá)HLA-A*02:01、HLA-A*02:07、HLA-A*11:01或HLA-A*24:02的靶細(xì)胞文庫來呈遞新冠病毒的抗原表位。在檢測了新冠病毒康復(fù)者的HLA-A基因型之后,將基因型匹配的靶細(xì)胞文庫和新冠病毒康復(fù)者的CD8 + T細(xì)胞進(jìn)行孵育,從涵蓋了上述四個(gè)HLA基因型的10個(gè)新冠病毒康復(fù)者的PBMC中分離了記憶性CD8+ T細(xì)胞進(jìn)行T-Scan篩選,共篩選到70條潛在的新冠病毒特異性CD8+ T細(xì)胞抗原識(shí)別表位,顯著富集了新冠病毒ORF1a、ORF1b和S蛋白的多肽片段,說明這三個(gè)基因可能是CD8+ T細(xì)胞介導(dǎo)的細(xì)胞免疫的重要抗原來源。
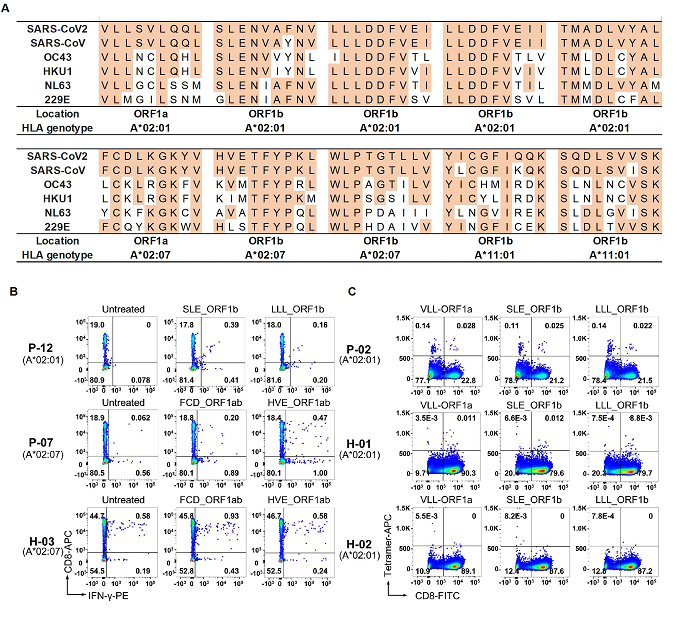
圖4: 保守性CD8+ T細(xì)胞抗原表位
作者首先報(bào)道了新冠病毒和其他冠狀病毒(包括SARS,和其他4種常見的引起普通感冒的冠狀病毒)在氨基酸序列上高度保守的抗原表位(圖4),并通過細(xì)胞內(nèi)INF-y染色的實(shí)驗(yàn)驗(yàn)證了其中7條多肽能夠在健康人中誘導(dǎo)CD8 + T細(xì)胞的免疫應(yīng)答,這也驗(yàn)證了先前對(duì)未感染新冠人群的預(yù)存的細(xì)胞免疫記憶的研究,表明人體內(nèi)如果有針對(duì)其它冠狀病毒的細(xì)胞免疫應(yīng)答,也會(huì)針對(duì)新冠病毒有一定的交叉免疫反應(yīng),而這種預(yù)存的細(xì)胞免疫的水平高低也可能部分決定了人群的病情差異。
新冠病毒的S蛋白介導(dǎo)病毒進(jìn)入細(xì)胞,是中和抗體的主要識(shí)別靶點(diǎn),S蛋白上的突變被證實(shí)能夠?qū)е虏《咎右葜泻涂贵w,也因此最為受到關(guān)注。圖5列出了4種關(guān)注變種的起源以及其在新冠病毒S蛋白上的突變位點(diǎn)。
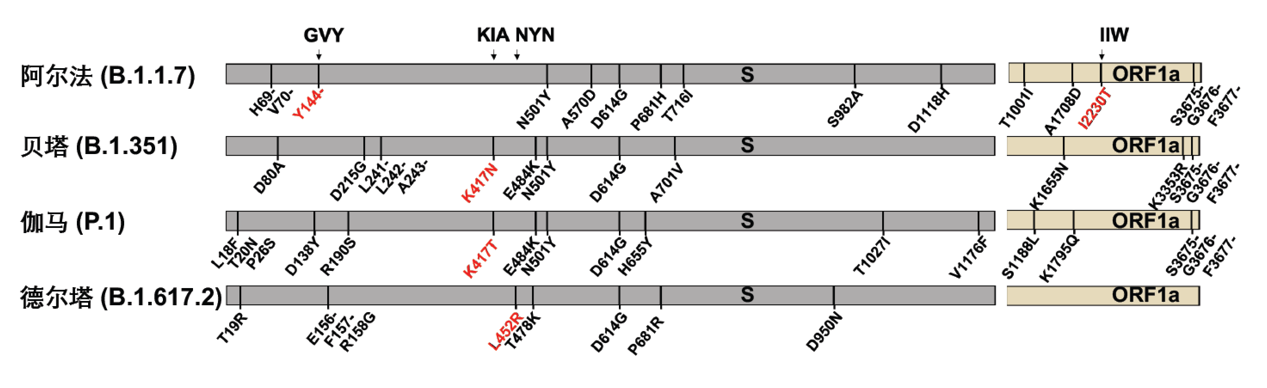
圖5: 新冠病毒的4種全球關(guān)注變種(關(guān)注變種)及其所攜帶的氨基酸突變位點(diǎn)(來自網(wǎng)站https://covariants.org )
為了探究關(guān)注變種的點(diǎn)突變是否幫助病毒逃逸CD8+ T細(xì)胞的免疫識(shí)別,作者重點(diǎn)關(guān)注了覆蓋關(guān)注變種突變位點(diǎn)的4條抗原表位: GVY、KIA、NYN、IIW;這四條抗原表位分別包括了S蛋白上的Y144-缺失突變、K417N/T突變、L452R突變和ORF1a蛋白上的I2230T突變。通過胞內(nèi)IFN-γ檢測和MHC-四聚體染色的方法,作者驗(yàn)證了這四條抗原表位能夠有效引起新冠康復(fù)患者或疫苗免疫后志愿者的CD8+ T細(xì)胞免疫應(yīng)答。為了進(jìn)一步探究關(guān)注變種攜帶的突變對(duì)CD8+ T細(xì)胞免疫的影響,作者合成了攜帶突變位點(diǎn)的抗原多肽,并比較了野生型抗原多肽和突變多肽的抗原有效性。對(duì)比發(fā)現(xiàn),和野生型多肽相比,新冠康復(fù)患者/疫苗免疫后志愿者的CD8+ T細(xì)胞對(duì)攜帶突變位點(diǎn)的抗原多肽的響應(yīng)能力下降(圖6)。這說明了這些位點(diǎn)的氨基酸對(duì)抗原識(shí)別及細(xì)胞免疫的產(chǎn)生非常重要,而關(guān)注變種將這些關(guān)鍵氨基酸突變之后,可能會(huì)降低病毒對(duì)細(xì)胞免疫的刺激,從而實(shí)現(xiàn)細(xì)胞免疫的逃逸。
圖6:抗原表位的突變降低了對(duì)疫苗接種者及新冠康復(fù)者體內(nèi)CD8+ T細(xì)胞激活的效率
更近一步,金騰川課題組通過晶體學(xué)的方法,解析了其中2條抗原表位和對(duì)應(yīng)MHC分子的復(fù)合物晶體結(jié)構(gòu)(圖7)。可以清晰地看到在KIA-HLA-A*02:01復(fù)合物(圖7A)中,417位的賴氨酸的正電荷側(cè)鏈與HLA-A*02:01蛋白的W167通過Pi陽離子相互作用牢牢結(jié)合,貝塔變種和伽馬變種攜帶的K417N/T突變將打破這一相互作用,導(dǎo)致HLA-A*02:01蛋白無法識(shí)別這一抗原表位,作者也通過生化的方法證明了這一結(jié)論。在NYN-HLA-A*24:02復(fù)合物(圖7B)中,452位的亮氨酸的疏水側(cè)鏈埋在了HLA-A*24:02蛋白表面的疏水口袋中,通過疏水相互作用緊密結(jié)合,關(guān)鍵位點(diǎn)452位氨基酸的突變可能幫助病毒逃逸抗原表位NYN介導(dǎo)的細(xì)胞免疫。
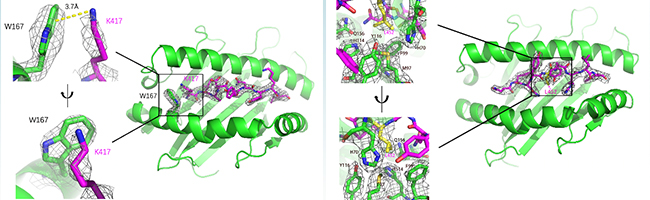
圖7: KIA&HLA-A*02:01復(fù)合物 (A) 和NYN&HLA-A*24:02復(fù)合物 (B) 的晶體結(jié)構(gòu)
以德爾塔變種為例,德爾塔變種攜帶的L452R的突變將引入帶正電荷的精氨酸R,完全破壞了原有的疏水相互作用,進(jìn)而破壞NYN抗原表位的呈遞。2021年7月Motozono等人在《細(xì)胞·宿主和微生物》(Cell Host & Microbe)上報(bào)道了L452R介導(dǎo)新冠病毒的細(xì)胞免疫逃逸,這篇研究也為此進(jìn)一步提供了結(jié)構(gòu)學(xué)基礎(chǔ)。
新冠病毒關(guān)注變種對(duì)免疫系統(tǒng)的逃逸大大增加了全球疫苗接種計(jì)劃的復(fù)雜度,了解關(guān)注變種免疫逃逸機(jī)制對(duì)未來疫苗的設(shè)計(jì)具有重要意義。先前的研究證實(shí)了HIV的進(jìn)化動(dòng)力之一就是通過突變CD8+ T細(xì)胞識(shí)別的抗原表位來達(dá)到逃逸細(xì)胞免疫的目的。這篇文章中,作者通過T-Scan技術(shù)篩選并鑒定了4條新冠病毒特異性CD8+ T細(xì)胞識(shí)別的關(guān)鍵性抗原表位,而新冠病毒關(guān)注變種(阿爾法、貝塔、伽馬和德爾塔)至少突變了其中一條關(guān)鍵性抗原表位,揭示了逃逸人體細(xì)胞免疫是新冠流行變種的普遍特點(diǎn)。除此之外,作者篩選到的冠狀病毒保守性抗原表位,也為后續(xù)的冠狀病毒的通用疫苗設(shè)計(jì)提供了理論基礎(chǔ)。
清華大學(xué)藥學(xué)院博士生張航和中國科學(xué)技術(shù)大學(xué)生命科學(xué)與醫(yī)學(xué)部博士生鄧莎莎為本文共同第一作者,譚旭教授和金騰川教授為本文共同通訊作者。該研究受到了國家自然科學(xué)基金委,清華大學(xué)春風(fēng)基金以及中科院先導(dǎo)項(xiàng)目的資助。清華大學(xué)董晨實(shí)驗(yàn)室、林欣實(shí)驗(yàn)室和石彥實(shí)驗(yàn)室為本項(xiàng)目提供了大力幫助。
論文鏈接:
https://www.cell.com/cell-reports/fulltext/S2211-1247(21)01155-4
供稿:藥學(xué)院
編輯:李華山
審核:呂婷